![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 47) Ano 2017. Pág. 13
Hallyson OLIVEIRA 1; Ronaldo do NASCIMENTO 2; Armindo Bezerra LEÃO 3; José Alberto Ferreira CARDOSO 4; Rafaela Félix Basílio GUIMARÃES 5
Recebido: 26/05/2017 • Aprovado: 25/06/2017
RESUMO: Esta pesquisa foi desenvolvida com o objetivo de avaliar os efeitos dos estresses hídrico e salino, induzidos por concentrações de Polietilenoglicol 6000 (PEG 6000) e NaCl, respectivamente, sobre a germinação de sementes e no crescimento inicial de plântulas de três variedades de algodão – BRS 286, BRS Topázio e BRS Verde. Os experimentos aconteceram em laboratório da Universidade Federal de Campina Grande – UFCG, onde as sementes foram semeadas em três folhas de papel Germitest. O delineamento adotado para cada experimento foi o inteiramente casualizado no esquema fatorial 3 x 5, com cinco repetições (25 sementes/repetição), totalizando 75 parcelas, sendo três variedades de algodão e cinco diferentes tratamentos: estresse salino – NaCl (0,02; 2,5; 5,0; 7,5; 10,0 dS m-1) e estresse hídrico – PEG 6000 (0,0; -0,3; -0,6; -0,9; -1,2 Mpa). Os crescentes níveis de CEa não afetaram a porcentagem de germinação, velocidade média e o índice de velocidade de germinação. Porém, o crescimento, as massas fresca total e seca total da plântula foram reduzidas à medida que os níveis de sais aumentaram, sendo o efeito maior na CEa de 10 dS m-1. O estresse hídrico induzido por concentrações de PEG 6000 causou mais danos à germinação e estabelecimento das plântulas, pois todas as variáveis estudadas foram afetadas pelos efeitos dos potenciais hídricos mais negativos, reduzindo-as acentuadamente a partir do potencial de -0,9 MPa, e não ocorrendo germinação no de -1,2 MPa. A variedade BRS 286 foi a mais resistente aos níveis de estresse hídrico e salino estudados, seguida da BRS Topázio e BRS Verde, respectivamente. |
ABSTRACT: This work was developed with the objective of evaluating the effects of water and saline stress, induced by concentrations of polyethylene glycol 6000 (PEG 6000) and NaCl, in seed germination and initial seedling growth of three cotton varieties – BRS 286, BRS Topazio e BRS Verde. The experiments were carried out at the laboratory of the Federal University of Campina Grande – UFCG, where the seeds were sown on three sheets of Germitest paper. The experimental design adopted for each experiment was completely randomized in the 3 x 5 factorial scheme, with five replicates (25 seeds/replicate), totaling 75 experimental plots, being three cotton varieties and five different treatments: saline stress – NaCl (0,02; 2,5; 5,0; 7,5; 10,0 dS m-1) and water stress – PEG 6000 (0,0; -0.3; -0,6; -0,9; -1,2 MPa). The increasing salt levels did not affect the percentage of germination, mean germination speed and germination velocity index, however, the seedling length, total fresh mass and total dry mass of the seedling was reduced as salt levels increased, with the major effect on the CEa of 10 dS m-1. The water stress induced by PEG 6000 concentrations caused more damage to germination and seedling establishment, because all the variables studied were affected by the effects of the most negative water potentials, reducing them significantly from the potential of -0,9 MPa, and no germination occurring at -1,2 MPa. The BRS 286 variety was the most resistant to the water and saline stress levels studied, followed by BRS Topazio and BRS Verde, respectively. |

As sementes são estruturas biológicas protegidas pelo tegumento, que é responsável por várias funções, dentre as quais, regula a germinação pela absorção de água na sua superfície, além disso, envolve o tecido de reserva cotiledonar e o eixo embrionário, este com seu crescimento temporariamente estável (Kijel & Galili, 1995). Quando umedecida em quantidade adequada, a semente inicia o processo de reativação ou crescimento do eixo embrionário, demonstrando a sua capacidade de produzir uma plântula normal, caracterizando, assim, o processo de germinação (Ferraz & Calvi, 2010).
A absorção de água ou embebição dá início a diversos processos físicos, fisiológicos e bioquímicos, os quais são prejudicadosna presença de fatores limitantes, dificultando assim a protrusão da radícula (Marcos Filho, 2005).
Entre os variados fatores limitantes que afetam o processo de germinação das sementes, destaca-se o estresse hídrico, pois a água é fundamental para o início desse processo, que faz parte direta ou indiretamente do desenvolvimento das outras fases da planta (Stefanello et al., 2006). A deficiência hídrica afeta a porcentagem, velocidade e a uniformidade de germinação (Filho, 2005).
O estresse salino é outro fator que possibilita problemas nas respostas de germinação das sementes, pois altos níveis de sais diminuem o potencial osmótico do substrato, o que dificulta a absorção de água pelas raízes. Além disso, causa toxidade no embrião, devido à presença de concentrações elevadas de íons (Prisco & O’Leary, 1970). Quando o potencial osmótico é bastante baixo, sendo inferior ao potencial osmótico das células do embrião, a porcentagem, a velocidade de germinação e a formação das plântulas são reduzidas (Marcos Filho, 2005).
Em laboratório, as fases de germinação e crescimento das plântulas têm sido bastante estudadas sob efeito desses estresses abióticos, por diversos autores (Souza et al., 2010; Pereira et al., 2011; Dutra, et al., 2014), principalmente da cultura do algodão (Ribeiro et al., 2002; Lima, et al., 2007; Queiroga, et al., 2008; Daniel et al., 2011; Barrocas, et al., 2014), que possui variedades tolerantes à seca e à salinidade, propiciando uma gama de resultados de elevada importância científica, servindo de base para o produtor rural na escolha da variedade, levando-se em consideração a quantidade e qualidade da água que dispõe para a produção. Tais estresses, em geral, são determinados no laboratório pelo uso de alguns compostos químicos, como o polietileno glicol 6000 (PEG 6000) e o cloreto de sódio (NaCl). O primeiro é utilizado na simulação do estresse hídrico, por ser inerte e não tóxico às sementes (Cordero & DI Stéfano 1991), simulando, de modo satisfatório, baixos potenciais de água, proporcionando à semente só a água, pois o mesmo não é absorvido (Villela et al., 1991). O segundo eleva o teor de sais no substrato, inibindo a germinação devido à diminuição do potencial osmótico, ocasionando prejuízos às demais fases do processo (Lima et al., 2005).
Portanto, essa pesquisa teve como objetivo avaliar os efeitos dos estresses hídrico e salino, induzidos por concentrações de PEG 6000 e NaCl, respectivamente, na germinação das sementes e no crescimento inicial de plântulas de algodão.
Os experimentos foram conduzidos em laboratório da Universidade Federal de Campina Grande – UFCG, Campus I. As sementes de algodão utilizadas no estudo foram a BRS Topázio, a BRS Verde e a BRS 286, doadas pela Embrapa Algodão, Campina Grande - PB.
As sementes passaram pelo processo de deslintamento químico via úmida, ou seja, a retirada do línter utilizando ácido sulfúrico, na proporção de 7 kg de sementes por litro de ácido. E, após o deslintamento, foram submetidas a uma lavagem com água e hidróxido de sódio (NaOH) a 5%, para a remoção dos resíduos de línter e ácido sulfúrico.
O delineamento experimental adotado para cada experimento foi o inteiramente casualizado no esquema fatorial 3 x 5, com cinco repetições (25 sementes/repetição), totalizando 75 parcelas experimentais, sendo três variedades de algodão herbáceo (BRS Topázio, BRS Verde e BRS 286) e cinco diferentes tratamentos: estresse salino – cloreto de sódio (0,02; 2,5; 5,0; 7,5; 10,0 dS m-1) e estresse hídrico – polietilenoglicol 6000 (0,0; -0,3; -0,6; -0,9; -1,2 Mpa).
As soluções de cloreto de sódio (NaCl) foram preparadas de acordo com a tabela proposta por Richards (1980) e as de polietilenoglicol 6000 (PEG 6000), conforme a tabela sugerida por Villela et al. (1991), à temperatura de 25 ºC.As sementes foram semeadas em três folhas de papel Germitest, utilizando duas folhas como base e uma para cobrir, e umedecidas com as soluções de NaCl e PEG 6000 em quantidade três vezes o peso do papel seco. Posteriormente, as folhas foram enroladas e colocadas em sacolas plásticas transparentes e vedadas para reduzir a perda de umidade. Os rolos, com as sementes, permaneceram em câmaras de germinação à temperatura de 25 ºC, por um período de 15 dias (Brasil, 2009). A avaliação do número de sementes germinadas ocorreu diariamente, adotando-se como critério para a leitura de semente germinada, a emissão de raiz (Brasil, 2009).
Os outros parâmetros avaliados foram:

Além do comprimento da plântula (CP), a massa fresca total da plântula (MFTP) e massa seca total da plântula (MSTP).
O comprimento das plântulas foi determinado com o auxílio de uma régua milimetrada. Elas secaram em estufa por um período de 24 horas à temperatura de 65 ºC, para determinação de MSTP.
Os dados obtidos foram submetidos à análise de variância e de regressão e teste de médias, sendo estas comparadas pelo teste de Tukey, a 5% de probabilidade (Ferreira, 2011).
De acordo com a Tabela 1, os diferentes níveis crescentes de salinidade (S) exerceram efeito significativo (p < 5%) apenas sobre o tempo médio de germinação (TMG), bem como não ocorreu interação significativa entre os fatores salinidade e genótipos (S x G), o que demonstra, possivelmente, que esses níveis de sais utilizados não afetam os processos fisiológicos e bioquímicos necessários, para proporcionar a germinação das sementes estudadas.
Apesar de corrente, o conhecimento de que a salinidade afeta o potencial osmótico da solução, e, consequentemente, o gradiente de potencial hídrico entre substrato e a semente, dificultando o processo de embebição e comprometendo a germinação (O'Leary, 1995), não foi observado no presente estudo efeito significativo para tal comportamento. Supõe-se que as cultivares avaliadas possuem uma tolerância aos efeitos produzidos pelo estresse salino nas suas germinações. Pereira (2012), ao estudar o estresse salino, em dez cultivares de algodão colorido, constatou que a variedade BRS Topázio apresentou uma tolerância maior, observando uma PG e um IVG satisfatórios, diferindo da cultivar BRS Verde, afetada na sua fase de germinação. No entanto, o autor atribui que, possivelmente, essa diferença não tenha sido ocasionada pelo efeito dos sais, mas sim, devido ao vigor das sementes. Em estudo realizado com variedades de algodão de fibra branca e colorida, submetidas a diferentes níveis de salinidade, Lopes & Silva (2010) constataram efeitos negativos em todas as variáveis da germinação analisadas, com maiores danos à germinação nos níveis a partir da CEa de 7,5 dS m-1.
Ao serem analisados os genótipos (G), verificou-se diferença significativa para as variáveis estudadas, a um nível de significância a 5% de probabilidade (Tabela 1).
Tabela 1: Resumo da Análise de Variância referente à porcentagem de germinação (PG),
tempo médio de germinação (TMG), velocidade média de germinação (VMG) e índice de
velocidade de germinação (IVG) de sementes de algodão, submetidas a diferentes
níveis de salinidade (NaCl).
Fator de Variação |
GL |
ANAVA - NaCl |
|||
PG |
TMG |
VMG |
IVG |
||
|
---%--- |
--dias-- |
-dias-1- |
|
|
Salinidade (S) |
4 |
ns |
* |
ns |
ns |
Genótipo (G) |
2 |
* |
* |
* |
* |
S x G |
8 |
ns |
ns |
ns |
ns |
Bloco |
4 |
ns |
ns |
ns |
ns |
CV (%) |
|
5,75 |
0,81 |
2,58 |
5,68 |
ns – não significativo; * - p < 5% pelo Teste de Tukey
Na Figura 1, estão apresentados os resultados médios obtidos das variáveis de porcentagem de germinação (PG), tempo médio de germinação (TMG), velocidade média de germinação (VMG) e índice de velocidade de germinação (IVG) referente ao fator genótipos.
A variedade que demonstrou melhores resultados, em relação à PG, TMG, VMG e IVG, foi a BRS 286, seguida da BRS Topázio, que diferiu significativamente da primeira somente nas variáveis TMG e VMG. A BRS Verde obteve menores valores médios, diferindo das outras cultivares para essas variáveis.
Os processos fisiológicos das sementes são caracterizados geneticamente durante sua formação. Além disso, as características genéticas dos descendentes se concretizam pela união dos gametas masculinos e femininos e são estabelecidas no momento da fecundação. Com isso, o desempenho das sementes na germinação varia entre espécies e cultivares (Filho, 2005).
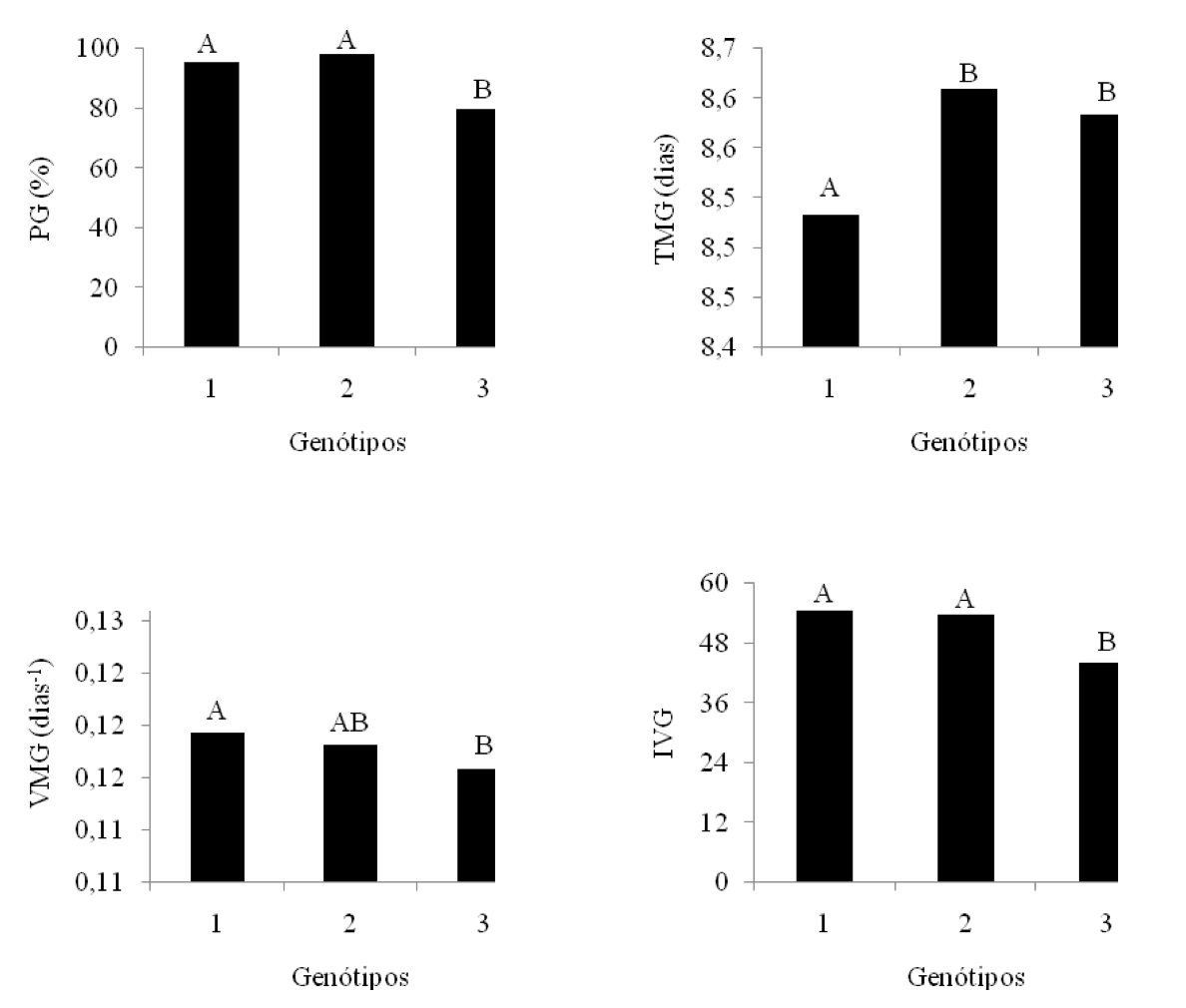
Figura 1
Valores médios das variáveis: porcentagem de germinação (PG),
tempo médio de germinação (TMG), velocidade média de
germinação (VMG) e índice de velocidade de germinação (IVG)
para o fator genótipos. Médias com a mesma letra não diferem
significativamente pelo Teste de Tukey, a 5% de probabilidade.
1 – BRS 286; 2 – BRS Topázio; 3 – BRS Verde.
Furtado et al. (2007) relatam que as diferentes características fisiológicas e morfológicas de cada cultivar de algodão proporcionam comportamentos distintos em cada uma quando submetidas aos efeitos da salinidade.
Constata-se, conforme os resultados obtidos da análise de variância (Tabela 2), que a salinidade afetou estatisticamente o crescimento (CP) e a massa seca total das plântulas (MSTP). E o fator genótipos apresentou diferença significativa nas variáveis de comprimento (CP) e massa fresca total das plântulas (MFTP), a um nível de significância de 5% de probabilidade, não se observando efeito na interação entre salinidade e genótipos (S x G).
Tabela 2
Resumo da Análise de Variância referente ao comprimento
da plântula (CP), massa fresca total da plântula (MFTP) e
massa seca total da plântula (MSTP) de cultivares de algodão
submetidas a diferentes níveis de salinidade (NaCl)
Fator de Variação |
GL |
ANAVA - NaCl |
||
CP |
MFTP |
MSTP |
||
---cm--- |
---g--- |
---g--- |
||
Salinidade (S) |
4 |
* |
ns |
* |
Genótipo (G) |
2 |
* |
* |
ns |
S x G |
8 |
ns |
ns |
ns |
Bloco |
4 |
ns |
ns |
ns |
CV (%) |
|
10,78 |
12,96 |
28,31 |
ns – não significativo; * - p < 5% pelo Teste de Tukey
Conforme os resultados da Figura 2A, relacionados ao comprimento da plântula (CP), a cultivar de fibra branca BRS 286 diferiu estatisticamente das outras, sendo a mais tolerante aos altos níveis de CEa (dS m-1), com um comprimento médio de 6,32cm, seguida das cultivares de fibra colorida BRS Verde (5,61cm) e BRS Topázio (5,23cm), que não diferenciaram entre si.
De acordo com a equação de regressão, a variável CP sofreu redução significativa linear, à medida que os níveis de salinidade aumentaram (Figura 2B). O melhor resultado ocorreu na CEa de 0,02 dS m-1, enquanto o menor crescimento aconteceu com a alta do nível de salinidade (10 dS m-1). Isso se deve, possivelmente, à maior dificuldade da plântula em absorver água, em maiores concentrações de sais solúveis no substrato.
De acordo com Dias & Blanco (2010), a planta reduz a absorção de água, em condições de estresse salino, devido ao aumento do potencial osmótico no substrato, que pode atingir um nível elevado, em que ela não terá força de sucção necessária para superar esse potencial. A acumulação excessiva de sais solúveis sobre as plantas dificulta a absorção de água, causa toxidez de íons específicos e interfere em seus processos fisiológicos, prejudicando, desse modo, seu crescimento (Dias et al., 2003).
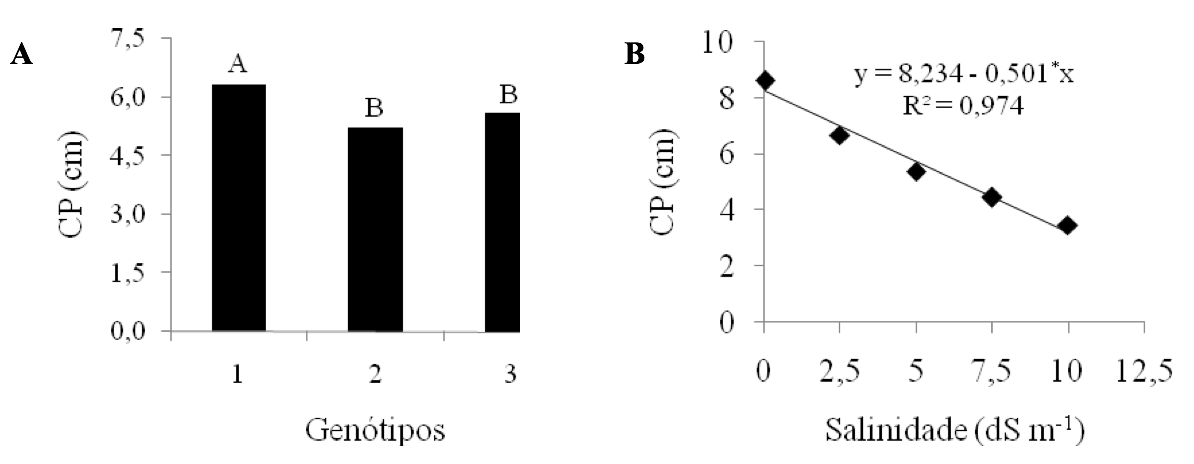
Figura 2A e 2B
Valores médios da variável comprimento da plântula (CP) para o fator genótipos
e salinidade, em função dos níveis crescentes de salinidade da água (dS m-1).
Médias com a mesma letra não diferem significativamente pelo Teste de Tukey a
5% de probabilidade. 1 – BRS 286; 2 – BRS Topázio; 3 – BRS Verde.
Para a variável massa fresca total da plântula (MFTP) houve diferença significativa entre genótipos (Figura 3), na qual a variedade BRS 286 obteve melhores resultados, seguida da BRS Topázio e BRS Verde. Tal aumento de massa, provavelmente, reflete em uma maior absorção de água pelas raízes, mesmo em condições de salinidade. Esses resultados contrariam os encontrados por Oliveira et al. (2013), que estudaram a tolerância das cultivares BRS Verde e BRS Topázio a diferentes níveis de salinidade da água de irrigação, verificando que os valores médios das massas frescas da raiz, caule e folhas não diferiram entre si, mediante as condições de salinidade estudadas.
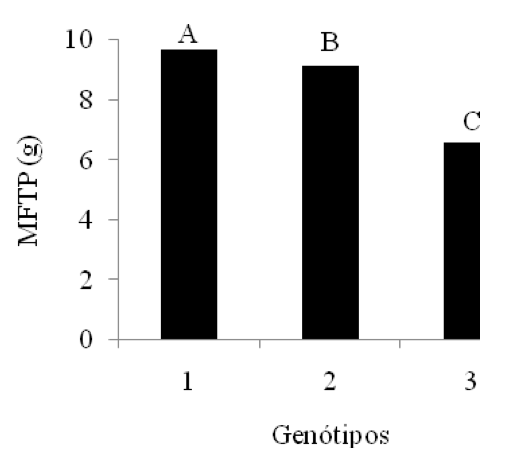
Figura 3: Valores médios da variável massa fresca total da plântula
(MFTP) para o fator genótipos. Médias com a mesma letra não
diferem significativamente pelo Teste de Tukey a 5% de probabilidade.
1 – BRS 286; 2 – BRS Topázio; 3 – BRS Verde.
Analisando a massa seca total da plântula (MSTP), em função da salinidade, verifica-se que os resultados médios obtidos reduziram à medida que os níveis de CEa aumentaram (Figura 4), em que na CEa de 0,02 dS m-1 foi observado maior massa seca (1,80g), e a menor, na CEa de 10,0 dS m-1 (0,89g). Resultados semelhantes foram encontrados por Lima (2007), que ao estudar a qualidade fisiológica de sementes de genótipos de algodão sob estresse salino, constatou que aos 20 dias após o início do teste de germinação, em rolos de papel Germitest, houve a diminuição da média de massa seca das plântulas de algodão, com o aumento do potencial salino.
A menor absorção de água pelas sementes reduz a velocidade dos processos fisiológicos e bioquímicos necessários para o estabelecimento de plântulas normais, apresentando menores comprimentos e menor acúmulo matéria seca (Sá, 2000).Ferreira et al. (2004) atribuem à redução do acúmulo de massa ao excesso dos íons Na+ e Cl-, que podem ser os responsáveis pela menor eficiência de atividade enzimática, ocasionando distúrbios na assimilação de nitrogênio e produção de energia.
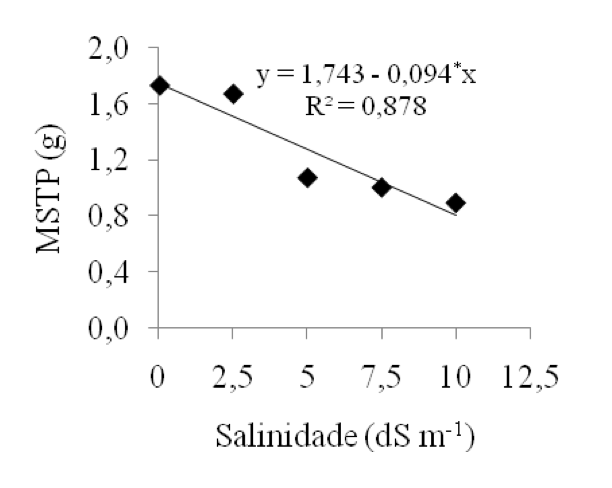
Figura 4: Massa seca total das plântulas, em função dos
níveis crescentes de salinidade da água (dS m-1).
Ventura (1956), estudando a linhagem 7888 de algodão “mocó”, verificou que as sementes submetidas à água destilada absorviam uma quantidade de água de 60,1% do seu peso inicial, contudo, quando expostas à solução de NaCl, essa absorção era de apenas 2,9%. Tal fato, provavelmente, pode evidenciar também a redução do crescimento das plântulas e de massa encontrados nesta pesquisa.
Conforme explícito na Tabela 3, todas as variáveis referentes à germinação sofreram efeito significativo (p < 5%) pelo fator potencial hídrico (P), o que evidencia, provavelmente, que as sementes de algodão reduziram sua capacidade germinativa quando expostas ao estresse hídrico. As variáveis porcentagem de germinação (PG) e índice de velocidade de germinação (IVG) também sofreram efeito significativo entre genótipos (G) e na interação (P x G).
Tabela 3: Resumo da Análise de Variância referente à porcentagem
de germinação (PG), tempo médio de germinação (TMG), velocidade média
de germinação (VMG) e índice de velocidade de germinação (IVG) de sementes
de algodão, submetidas a diferentes potenciais hídricos (PEG 6000).
Fator de Variação |
GL |
ANAVA – PEG 6000 |
|||
PG |
TMG |
VMG |
IVG |
||
|
---%--- |
--dias-- |
-dias-1- |
|
|
P. Hídrico (P) |
4 |
* |
* |
* |
* |
Genótipo (G) |
2 |
* |
ns |
ns |
* |
P x G |
8 |
* |
ns |
ns |
* |
Bloco |
4 |
ns |
ns |
ns |
ns |
CV (%) |
|
10,92 |
28,80 |
20,82 |
9,85 |
ns – não significativo; * - p < 5% pelo Teste de Tukey
Verifica-se na Figura 5 que o modelo de regressão que melhor se ajustou aos resultados médios obtidos, para as variáveis PG, TMG, VMG e IVG, foi o quadrático, apresentando um aumento dos valores dessas variáveis até o potencial hídrico (ψw) de -0,3 MPa, reduzindo significativamente a partir do ψw de -0,6 MPa.
A porcentagem de germinação (PG) e o índice de velocidade de germinação (IVG), que é calculado para estabelecer as diferenças na velocidade de germinação de grupos ou lotes de sementes (Brasil, 2009), reduziram, de maneira drástica, a partir do potencial hídrico de -0,6 MPa, não ocorrendo germinação no ψw de -1,2 Mpa. Demonstrando, provavelmente, que potencias hídricos muito próximos ao ponto de murcha permanente (-1,5 MPa) prejudicam de forma efetiva a capacidade germinativa e velocidade de germinação das sementes de algodão. Schlichting (2012) define ponto de murcha permanente como a quantidade de água retida no substrato que a planta não consegue mais retirar.
Outro fato que pode, possivelmente, explicar essa redução drástica de PG e IVG é a baixa solubilidade do PEG 6000 em água, que torna a solução altamente viscosa, dificultando assim a absorção de água pelas sementes. Munns (2002) afirma que sais de alta solubilidade, ou seja, menor peso molecular proporciona pequeno efeito sobre a “seca fisiológica” durante a germinação, em comparação com o PEG 6000. E com isso, as sementes, além de absorverem água, também absorvem esses sais solúveis, os quais proporcionam diminuição do potencial osmótico intracelular, e consequentemente, regulam a hidratação e turgescência celular, devido à manutenção do potencial hídrico entre a semente e o substrato. Em contrapartida, o PEG 6000, por ter um elevado peso molecular, não consegue penetrar nas células das sementes, pois suas moléculas são maiores, não atravessando as membranas celulares, possibilitando às sementes apenas a absorção de água, ocorrendo assim um maior estresse hídrico, quando utilizado em elevadas concentrações (Filho, 2005). Tais citações ainda podem possivelmente evidenciar a diferença de efeito significativo exercido pelos fatores salinidade (S) e potenciais hídricos (P) sobre a porcentagem de germinação das sementes (PG) e o índice de velocidade germinação (IVG) nesta pesquisa (Tabela 1 e 3).
Meneses (2007), estudando a qualidade fisiológica de sementes de diferentes variedades de algodão, submetidas ao estresse hídrico induzido por PEG 6000, encontrou resultados semelhantes a esta pesquisa relacionados à variável PG e IVG, em que os valores médios de porcentagem de germinação e velocidade de germinação reduziram drasticamente, a partir do potencial hídrico de -0,4 MPa, enquanto que no ψw de -1,0 MPa isso ocorreu quase por completo.
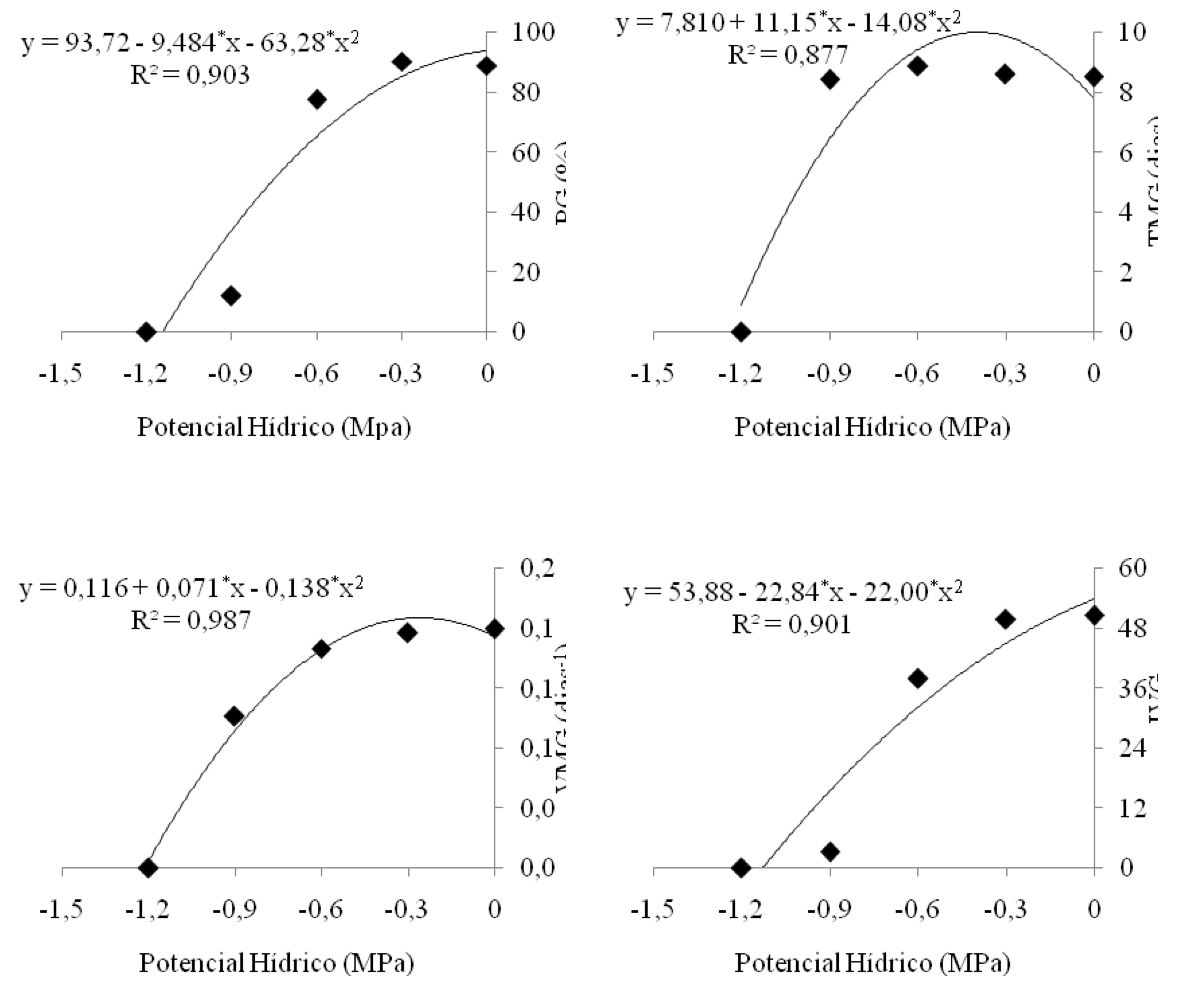
Figura 5: Porcentagem de germinação, tempo médio de germinação, velocidade
média de germinação e índice de velocidade de germinação, em função de diferentes
potenciais hídricos (MPa).
Diferente dos valores médios de PG e IVG, em função dos potenciais hídricos, o tempo médio de germinação (TMG) só apresentou decréscimo no ψw de -1,2 MPa, justamente no que não houve germinação (Figura 5). Nos demais potencias, os valores médios de TMG foram praticamente iguais, variando entre 8,61 e 8,44 dias, sendo provável que todas as sementes germinadas emitiram radículas praticamente no mesmo dia. Porém, a velocidade média de germinação (VMG) diminuiu efetivamente a partir do ψw de -0,6 (MPa), demonstrando que elas sofreram um retardo maior no início da germinação, com esse potencial hídrico.
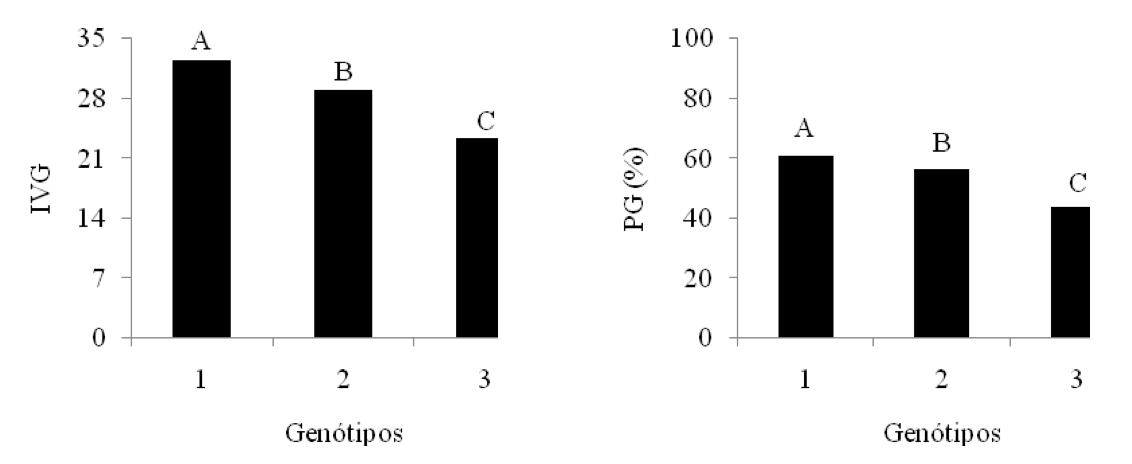
Figura 6: Valores médios das variáveis: porcentagem de germinação (PG) e índice
de velocidade de germinação (IVG) para o fator genótipos. Médias com a mesma
letra não diferem significativamente. 1 – BRS 286; 2 – BRS Topázio; 3 – BRS Verde.
As três cultivares estudadas diferiram estatisticamente entre si (p < 5%) com relação às variáveis PG e IVG (Figura 6), em que as sementes da BRS 286 obtiveram melhores resultados de germinação e maior velocidade em germinar, seguida da BRS Topázio e BRS Verde, respectivamente.
3.2.2. Comprimento, Massa Fresca e Massa Seca da Plântula
Os baixos níveis de potenciais hídricos exerceram efeito significativo (p < 5%) sobre o comprimento (CP), a massa fresca (MFTP) e seca total (MSTP) da plântula, comprovando que quanto menos disponibilidade de água para as raízes, menor o crescimento dela. Os fatores genótipo (G) e interação (P x G) estabeleceram efeito significativo (p < 5%) nas variáveis MFTP e MSTP (Tabela 4).
Tabela 4: Resumo da Análise de Variância referente ao comprimento da
plântula (CP), massa fresca total da plântula (MFTP) e massa seca total da
plântula (MSTP) de cultivares de algodão submetidas a diferentes potenciais
hídricos (PEG 6000).
Fator de Variação |
GL |
ANAVA – PEG 6000 |
||
CP |
MFTP |
MSTP |
||
cm |
g |
g |
||
P. Hídrico (P) |
4 |
* |
* |
* |
Genótipo (G) |
2 |
ns |
* |
* |
P x G |
8 |
ns |
* |
* |
Bloco |
4 |
ns |
ns |
ns |
CV (%) |
|
36,43 |
22,27 |
33,39 |
ns – não significativo; * - p < 5% pelo Teste de Tukey
O modelo de regressão que melhor se ajustou aos resultados médios obtidos de CP e MFTP foi o linear, resultando num decréscimo em função da redução do potencial hídrico no substrato, ou seja, da menor disponibilidade de água (Figura 7). Segundo Bewley & Black (1994), a indisponibilidade de água para a semente acarreta problemas no seu metabolismo, como reduções na digestão e translocação das reservas para as diversas partes da nova plântula. Durante a germinação, as reservas são principalmente constituídas de carboidratos, proteínas e lipídios, em que suas degradações dependem de várias enzimas hidrolíticas (Lacerda et al., 2007). As enzimas hidrolíticas ou digestivas só atuam em reações químicas na presença de moléculas de água, o que evidencia, provavelmente, que a falta de água para a semente afeta direto os processos bioquímicos relacionados à germinação, ocasionando posteriormente, reduções no crescimento e massa fresca da plântula.
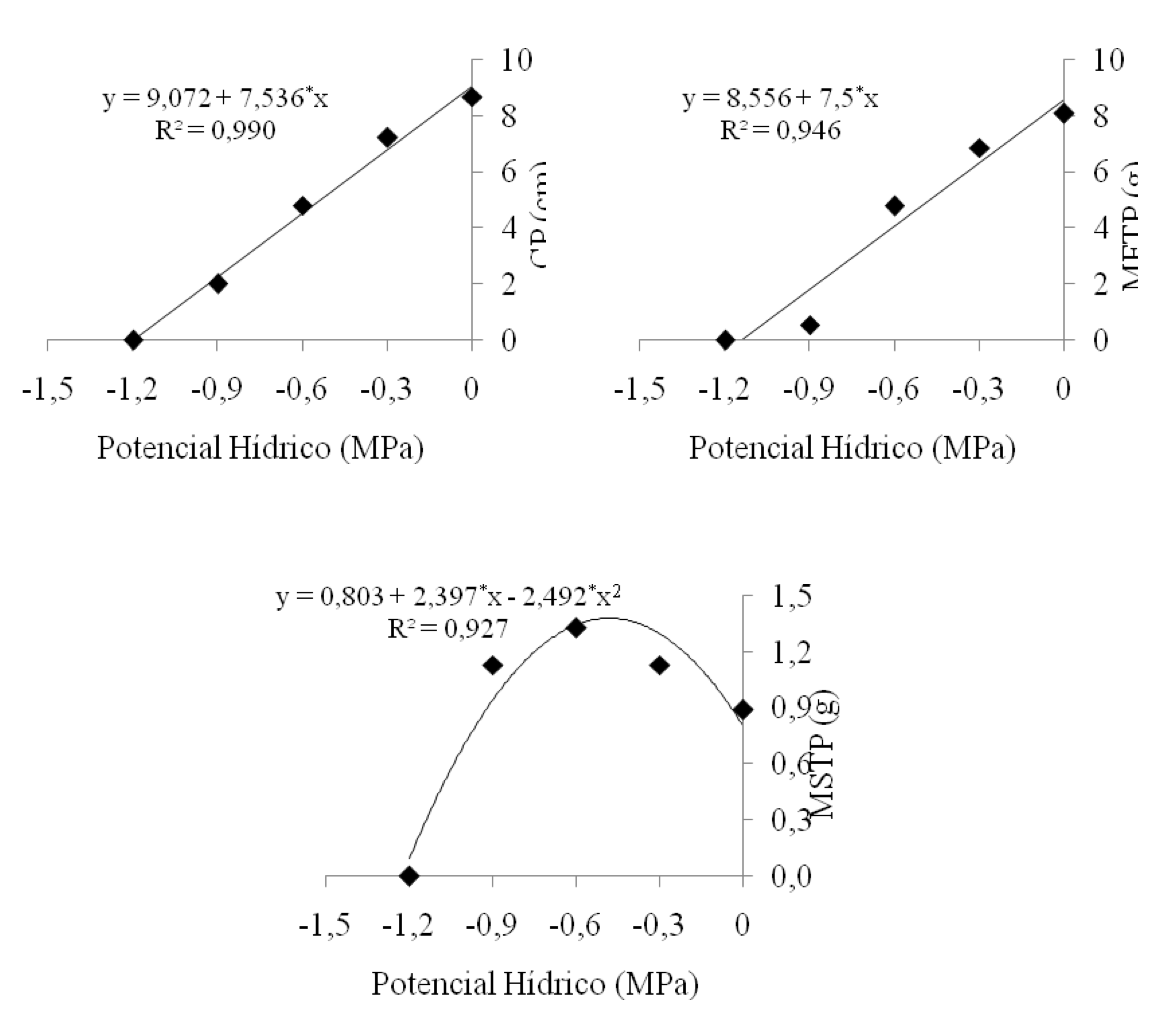
Figura 7: Comprimento da plântula (CP), massa fresca total da plântula
(MFTP) e massa seca total da plântula (MSTP), em função dos diferentes
potenciais hídricos (MPa).
Houve incremento de matéria seca até o potencial hídrico de -0,6 MPa, diminuindo de maneira drástica a partir ψw de -0,9 MPa (Figura 7). Esse incremento de biomassa seca pode estar relacionado a algum tipo de mecanismo de tolerância ao estresse hídrico, mantendo o vigor das plântulas. Távora & Melo (1991) afirmam que, dentre os mecanismos de tolerância ao estresse hídrico, os principais são o aumento e aprofundamento das raízes, podendo ser pelo alongamento das células ou ganho de massa seca, sendo este relacionado ao crescimento e aumento da espessura das paredes celulares (enrijecimento). Carneiro et al. (2011), analisando a atividade antioxidante e viabilidade de sementes de girassol, após aplicação de PEG 6000 e NaCl, verificaram que o PEG 6000 proporcionou um incremento de matéria seca até o potencial de -0,4 Mpa, tanto na parte aérea quanto nas raízes, e possibilitou um decréscimo drástico no potencial de -0,8 MPa.
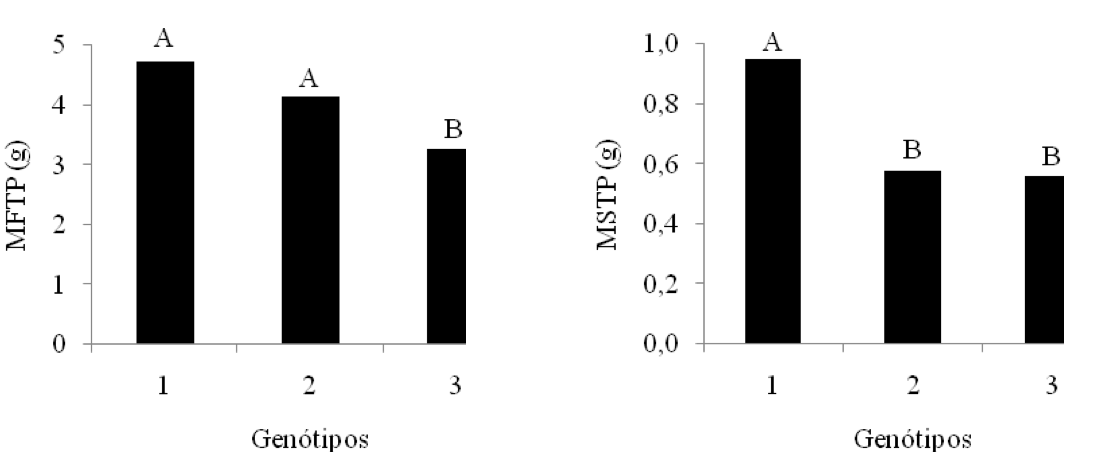
Figura 8: Valores médios das variáveis: massa fresca total da plântula (MFTP)
e massa seca total da plântula (MSTP) para o fator genótipos. Médias com
a mesma
letra não diferem significativamente. 1 – BRS 286; 2 – BRS Topázio; 3 – BRS Verde.
Conforme a Figura 8, a variedade de algodão BRS 286 apresentou melhores resultados de biomassa fresca (4,73 g) e seca (0,95 g), em relação às outras variedades estudadas, o que demonstra ser a cultivar com maior vigor, conseguindo, provavelmente, absorver mais água e tolerar os baixos níveis de disponibilidade de água no substrato.
A variedade que demonstrou maior tolerância aos estresses salino e hídrico, em todas as variáveis analisadas, foi a BRS 286, seguida da BRS Topázio e BRS Verde, respectivamente.
Os altos níveis de salinidade no substrato causaram redução significativa no vigor das sementes, sendo o de 10 dS m-1 o que causou mais decréscimo.
As sementes apresentaram tolerância ao estresse hídrico até o potencial hídrico de -0,6 Mpa, em todas as variáveis, reduzindo significativamente a partir de -0,9 Mpa.
As sementes sofreram problema efetivo de germinação no potencial hídrico de -1,2 MPa, não acontecendo emissão da radícula em nenhuma semente das variedades estudadas.
Barrocas, E. N., Machado, J. da C., Alves, M. de C., y Corrêa, C. L. (2014). Desempenho de sementes de algodão submetidas à deficiência hídrica e presença de Colletotrichum gossypii var. cephalosporioides. Bioscience Journal, 30(2), 421-428.
Bewley, J. D., Black, M. (1994). Seeds: physiology of development and germination. New York: Plenum Press, 2 ed. p. 445.
BRASIL. (2009). Regras para análise de sementes. Brasília: Ministério Da Agricultura, Pecuária e Abastecimento. p. 395.
Carneiro, M. M. L. C., Deuner, S., Oliveira, P. V. de., Teixeira, S. B., Sousa, C. P., Bacarin, M. A., y Moraes, D. M. de. (2011). Atividade antioxidante e viabilidade de sementes de girassol após estresse hídrico e salino. Revista Brasileira de Sementes, 33(4), 752-761.
Cordero, R. A. S., y DI Stéfano, J. F. G. (1991). Efecto dei estrés osmótico sobre la germinación de sernillas de Tecoma stans (Bignolliaceae). Revista Biologia Tropical, 39(1), 107-110.
Daniel, V. C., Sevilha, R. R., Silva, F. F. da., y Zonetti, P. da C. (2011). Germinação e crescimento de plântulas de algodão colorido sob condições de estresse salino. Revista em Agronegócios e Meio Ambiente, 4(2), 321-333.
Dias, N. D., y Blanco, F. F. (2010). Efeitos dos sais no solo e na planta. In: Gheyi, H. R., Dias, N. S., y Lacerda, C. F. Manejo da salinidade na agricultura: Estudos básicos e aplicados. Fortaleza-CE: Instituto Nacional de Ciência e Tecnologia em Salinidade. p. 129-140.
Dias, N. da S., Gheyi, H. R., Duarte, S. N. (2003). Prevenção, manejo e recuperação dos solos afetados por sais. Piracicaba: ESALQ/USP/LER. p. 118.
Dutra, T. R., Massad, M. D., Matos, P. S., Oliveira, J. C. de., y Sarmento, M. F. Q. (2014). Germinação e crescimento inicial de plântulas de carobinha-do-campo submetido ao estresse hídrico e salino. Agropecuária Científica no Semiárido, 10(4), 39-45.
Ferraz, I. D. K., y Calvi, G. P. (2010). Teste de germinação. In: Manual de Análise de Sementes. Manaus: UFAM. p. 55-122.
Ferreira, A. G., Borguetti, F. (2004). Germinação: do básico ao aplicado. Porto Alegre: Artmed. p. 324.
Ferreira, D. F. (2011). Sisvar: a computer statistical analysis system. Ciência e Agrotecnologia, 35(6), 1039-1042.
Filho, J. M. (2005). Fisiologia de sementes de plantas cultivadas. Piracicaba: Fealq. p. 495.
Furtado, R. F., Mano, A. R. O., Alves, C. R., Freitas, S. M., y Filho, S. M. (2007). Efeito da salinidade na germinação de sementes de algodão. Revista Ciência Agronômica, 38(2), 224-227.
Kijel, J., Galili, G. E. (1995). Seed development and germination. New York, Marcel Dekker Inc. p. 853.
Laboriau, L. G., y Valadares, M. E. B. (1976). On the germination of seeds Calostropisprocera (Ait.) f. Anais da Academia Brasileira de Ciências, 48(2), 263-284.
Lacerda, C. F. de, Filho, J. E., Pinheiro, C. B. (2007). Fisiologia Vegetal. Fortaleza: Universidade Federal do Ceará. 1 ed. p. 353.
Lima, L. H. G. de M. (2007). Qualidade fisiológica de sementes de genótipos de algodoeiro sob estresse salino. (Dissertação de Mestrado). Universidade Federal do Rio Grande do Norte. Natal. p. 36.
Lima, L. H. G. de M., Meneses, C. H. S. G., Lima, M. M. de A., Pereira, W. E., Fernades, P. D., Bruno, R. de L. A., y Vidal, M. S. Atributos fisiológicos de sementes de algodoeiro submetidas a estresse salino. Revista Brasileira de Oleaginosas e Fibrosas, 11(3), 173-184.
Lima, M. G. S., Lopes, N. F., Moraes, D. M., y Abreu, C. M. (2005). Qualidade fisiológica de sementes de arroz submetidas a estresse salino. Revista Brasileira de Sementes, 27(1), 54-61.
Lopes, K. P., y Silva. M. (2010). Salinidade na germinação de sementes de algodão colorido. Revista Verde de Agroecologia e Desenvolvimento Sustentável, 5(3), 274–279.
Maguire, J. D. (1962). Speed of germination-aid in selection and evaluation for seedling emergence and vigor. Crop Science, 2(1), 176-177.
Marcos Filho, J. (2005). Fisiologia de sementes de plantas cultivadas. Piracicaba: FEALQ. p. 495.
Meneses, C. H. S. G. (2007). Qualidade fisiológica de sementes de algodão submetidas a estresse hídrico induzido por polietilenoglicol-6000. (Dissertação de Mestrado). Universidade Federal da Paraíba. Areia. p. 50-54.
Munns, R. (2002). Comparative physiology of salt and water stress. Plant Cell Environment, 25(2), 239-250.
O'Leary, J. W. (1995). Adaptive components of salt tolerance. In: Pessarakli, M. (Ed.) Handbook of plant and crop physiology. New York: Marcel Dekker, Inc. p. 577-585.
Oliveira, L. L. P., Dias, N. S., Farias, W. C., Medeiros, L. C., y Ferreira, L. L. (2013). Tolerância de cultivares de algodão (Gossypium hirsutum) à salinidade da água de irrigação. Revista Verde de Agroecologia e Desenvolvimento Sustentável, 8(4), 232 - 237.
Pereira, E. R. de L. (2012). Tolerância de genótipos do algodão colorido ao estresse salino. (Dissertação de Mestrado). Universidade Estadual da Paraíba. Campina Grande. p. 37.
Pereira, M. D., y Lopes, J. D. (2011). Germinação e desenvolvimento de plântulas de pinhão manso sob condições de estresse hídrico simulado. Semina: Ciências Agrárias, 32(1), 1837-1842.
Prisco, J. T., y O’Leary, J. W. (1970). Osmotic and toxic effects of salinity on germination of Phaseolus vulgaris L. seeds. Turrialba, 20(1), 177-184.
Queiroga, V. P., Duran, J. M., Bruno, R. L. A., Santos, J. W., y Queiroga, D. A. N. (2008). Qualidade de sementes de algodão submetidas ao condicionamento osmótico e hídrico. Revista Caatinga, 21(2), 156-164.
Ribeiro, U. P., Pinho, E. V. de R. V., Guimarães, R. M., y Viana, L. de S. (2002). Determinação do potencial osmótico e do período de embebição utilizados no condicionamento fisiológico de sementes de algodão. Ciência e agrotecnologia, 26(5), 911-917.
Richards, L. A. (1980). Suelos Salinos y Sodicos. Instituto Nacional de Investigaciones Agrícolas. México. p. 171.
Sá, M. E. (2000). Relações entre qualidade fisiológica, disponibilidade hídrica e desempenho de sementes de soja (Glycine max (L.) Merrill). (Tese de Doutorado). Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo. Piracicaba. p. 174.
Schlichting, A. F. (2012). Cultura do milho submetida a tensões de água no solo e doses de nitrogênio. (Dissertação de Mestrado). Universidade Federal de Mato Grosso. Rondonopólis. p. 19.
Souza, Y. A., Pereira, A. L., Silva, F. F. S. da., Reis, R. C. R., Evangelista, M. R. V., Castro, R. D. de., y Dantas, B. F. (2010). Efeito da salinidade na germinação de sementes e no crescimento inicial de mudas de pinhão-manso. Revista Brasileira de Sementes, 32(2), 83-92.
Stefanello, R., Garcia, D. C., Meneses, N. L., y Wrasse, C. F. (2006). Influência da luz, temperatura e estresse hídrico na germinação e no vigor de sementes de anis. Revista Brasileira de Agrociência, 12(1), 45-50.
Távora, F. J. A. F., y Melo, F. I. (1991). Respostas de cultivares de amendoim a ciclos de deficiência hídrica: Crescimento vegetativo, reprodutivo e relações hídricas. Revista Ciência Agronômica, 22(01/02), 47-60.
Ventura, M. M. (1956). Absorção de água, equilíbrios higroscópicos e atividade respiratória em sementes de algodão “mocó” (Gossypium hirsutum L. var. Marie galante Hutch). Phyton, 7(1), 7-13.
Villela, F. A., Doni-Filho, L., y Sequeira, E. L. (1991). Tabela de potencial osmótico em função da concentração de polietileno glicol 6000 e da temperatura. Pesquisa Agropecuária Brasileira, 26(11/12), 1957-1968.
1. Unidade Acedêmica de Engenharia Agrícola. Universidade Federal de Campina Grande. hallysonolivera_@hotmail.com
2. Unidade Acedêmica de Engenharia Agrícola. Universidade Federal de Campina Grande. ronaldon453@gmail.com
3. Unidade Acedêmica de Engenharia Agrícola. Universidade Federal de Campina Grande. armindoleao@yahoo.com.br
4. Unidade Acedêmica de Engenharia Agrícola. Universidade Federal de Campina Grande. jalbertofcardoso@gmail.com
5. Unidade Acedêmica de Engenharia Agrícola. Universidade Federal de Campina Grande. rafaellafelix_@hotmail.com